The poster child of scientific theory.
 A panel depicting Charles Darwin, Turin Museum of Human Anatomy. Image: Commons.
A panel depicting Charles Darwin, Turin Museum of Human Anatomy. Image: Commons.
When On the Origin of Species was first published in 1859, to say it caused a scientific and political earthquake would be a vast understatement. The ideas put forward would become the basis of every biological field in existence today.
The foundations of the theory are observations that were made while Charles Darwin and Alfred Russel Wallace travelled the world, but since then there have been many expansions and breakthroughs. I?ve allocated these topics of evidence into 5 categories and I?ll do my best to explain the significance of each. But to start, a bit about natural selection.
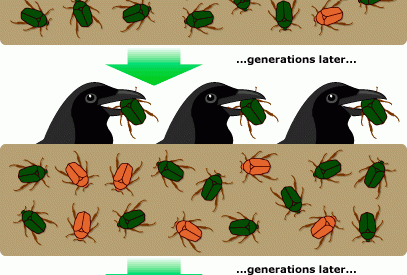
The 4 statements of evolution by natural selection.
 Image: University of California Museum of Paleontology?s Understanding Evolution.
Image: University of California Museum of Paleontology?s Understanding Evolution.
Evolution (which Darwin referred to as ?descent with modification?) is a multi-faceted affair, but the core mechanism proposed, known as natural selection, stands true to this day. It claimed four simple but powerful statements supported by decades of accumulated evidence:
- Variation in traits (or characteristics) within a species exists.
- These traits are heritable.
- Some traits result in individuals having more offspring (or are associated with it).
- These traits will increase within a population over time.
As such, natural selection is all about reproduction. Although Darwin was unaware of how variation come into existence (genetics was still some way off), it was only necessary for the variation to exist and for it to be heritable.
There are of course some notable exceptions to the above. As an example, a volcanic eruption, or meteor strike, can destroy an entire group of variants independent of their competitive ability or reproductive success (by chance). This is known as genetic drift, and it?s also an important mechanism that drives evolution.
The Evidence:
1. Biogeography.
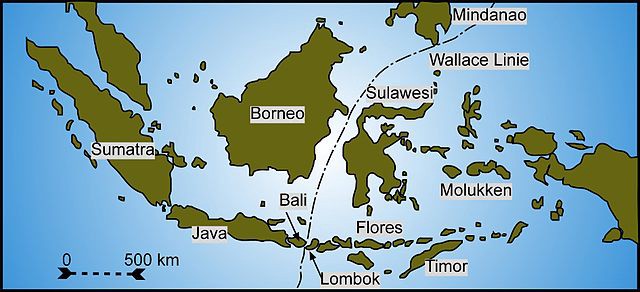 The ?Wallace Line?. During his extensive travels in Indonesia, Wallace noticed a stark difference between the species in the West and East, despite similar climate and terrain. Image: Gunnar Ries.
The ?Wallace Line?. During his extensive travels in Indonesia, Wallace noticed a stark difference between the species in the West and East, despite similar climate and terrain. Image: Gunnar Ries.
The modern field of biogeography revolves around the study of species distribution across the planet, and why such a distribution should exist. Wallace, arguably one of the founders of biogeography, noticed that species in one region were often very different from species in another, even though their climate and geography were the same. This didn?t fit with the conventional 1840s viewpoint of creation for purpose, and soon enough Wallace found many other examples which begged explanation, most famously the ?Wallace Line?.
Even though there was a very short distance between them, the species of eastern Indonesia (and Papua New Guinea) are very different from their counterparts in western Indonesia. The eastern species resemble those of the Australian continent, whilst the western species are more Asian. Wallace was able to draw a line between these two groups (see Wallace Line above).
We now know the differences are due to colonisation from the two different continents, with the various species later becoming isolated by rising waters ? adapting in isolation and becoming separate species. Colonised volcanic islands follow the same trend: Darwin?s Galapagos finches, isolated far into the Pacific, clearly had a common ancestor on the South/Central American mainland.
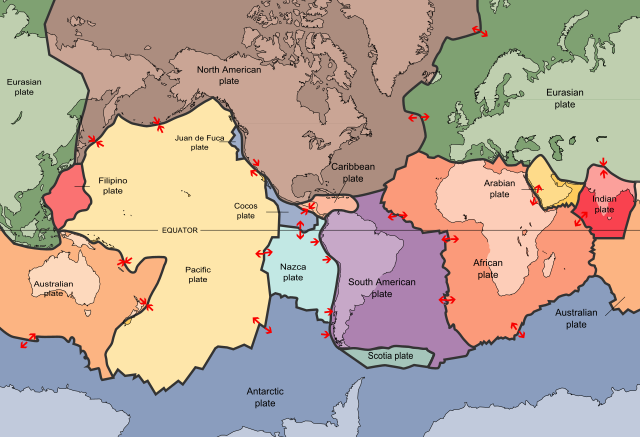 Global tectonic plates ? you can see the Wallace-line loosely represented between the Eurasian and Australian plates. Antarctica, South America, Africa, India, and Australasia were all once part of a mega-continent: Gondwana. (Image: USGS)
Global tectonic plates ? you can see the Wallace-line loosely represented between the Eurasian and Australian plates. Antarctica, South America, Africa, India, and Australasia were all once part of a mega-continent: Gondwana. (Image: USGS)
Following Alfred Wegener?s plate tectonic theory in the early 20th century (shown above), it became clear that inter-continental similarities between species could be explained by the movement, and breaking, of mega-continents. Indeed, one of the most convincing pieces of evidence behind the theory was that identical fossil plants and animals had been discovered on opposite sides of the Atlantic.
2. The Fossil Record.
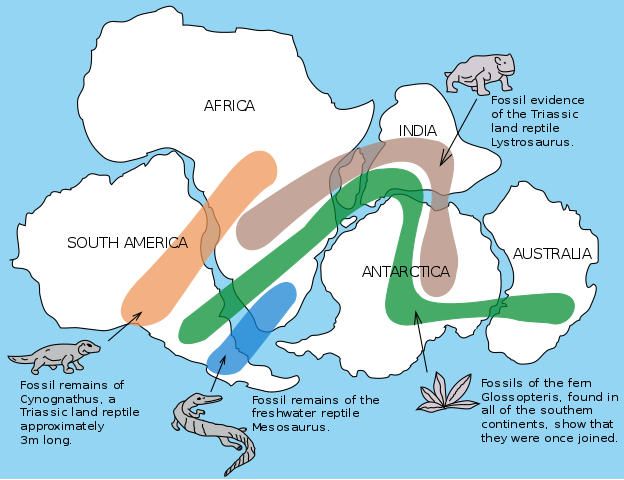 A graphic of Gondwana. Continental drift explains the distribution of the fossil record, and why identical fossils can be found on 3 different continents, separated by thousands of kilometres of ocean. Image: USGS.
A graphic of Gondwana. Continental drift explains the distribution of the fossil record, and why identical fossils can be found on 3 different continents, separated by thousands of kilometres of ocean. Image: USGS.
Changes in species distribution over time are very clear in the fossil record; as the dating of a fossil, or at the very least its surroundings, is possible*. This allows for a timestamp that gives us a tiny snapshot of what the species distribution looked like at certain periods in our planet?s history. The record is highly incomplete for many lineages, due to the rare conditions required for fossil formation, but I?ll focus on the fairly complete lineage of the birds as an example.
*Dating of geological specimens is not just limited to radiocarbon, which has a range of only around 70000 years. There are a variety of other methods, including K-Ar and Uranium-Lead dating, which both have ranges in the billions of years. It is pivotal to understand the vast geological timelines that facilitate evolution, as this is a common sticking point. Read more about the dating of rocks and fossils here.
The fossil record provides us with some inkling of a mass extinction event that wiped out many species some 66 million years ago. The only line of dinosaurs that survived, a small branch of the archosaurs, are the ancestors that led to modern birds.
 An Archaeopteryx fossil, an intermediary species between modern birds and dinosaurs. Image: James L. Amos.
An Archaeopteryx fossil, an intermediary species between modern birds and dinosaurs. Image: James L. Amos.
The first Archaeopteryx fossil was discovered in 1860, and although it looked like a bird, there were some distinct differences (see above). It had feathers, but it also had a bony tail and teeth ? both of which aren?t found in modern bird species. The bone structure was also somewhat different. All of this pointed to Archaeopteryx as an intermediary between birds and an earlier dinosaur.
The bone structure and presence of feathers in the fossils of theropods, a ground-dwelling dinosaur, suggest that they were the intermediary between Archaeopteryx and larger, pre-feather dinosaurs (although where the dinosaurs branched into feathered and non-feathered is contentious). As a result, using phylogenies (a family tree for relatedness over time), researchers have been able to piece together the legacy of the dinosaurs.
A complete fossil record will probably never be achieved, for fairly obvious reasons. However, great strides have been made in tracing other lineages: especially our own Homo genus. The fossil record is indicative of evolution, but it is only one piece in the evidence puzzle.
3. Embryology, similarity, and vestigial structures.
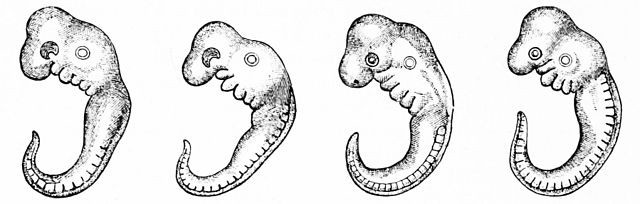 Embryos of four different species. From Left to Right; chicken, tortoise, pig, human. Image: Adapted from T. Bruntun.
Embryos of four different species. From Left to Right; chicken, tortoise, pig, human. Image: Adapted from T. Bruntun.
Many species share an embryonic larval structure (see above). Darwin postulated that: ?community of embryonic structure reveals community of descent?, and there?s a reason for this.
We can view the larvae, or embryos, as a template for shared ancestry. For instance, both human and fish embryos have gill-slits ? but these slits are removed in the later development of the human embryo, whilst developing into gills in fish. Genetic factors, developed through adaptation to land-environments, silence gill development in humans; but the early, gill-slit stage suggests we share a common ancestor with fish. Other examples include eyes in embryonic moles and teeth in embryonic whales.
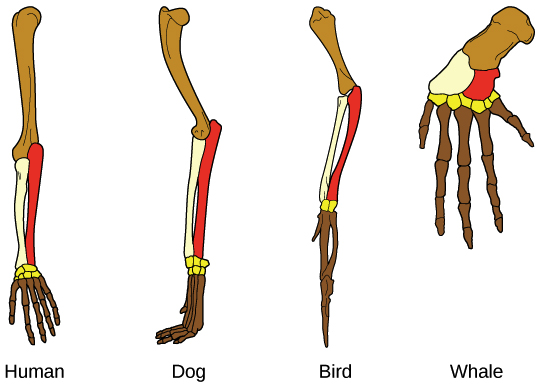 Homologous structures of the limb. Homology refers to features that are shared among different species due to descent from a common ancestor. Image: Understanding Evolution, Figure 7.
Homologous structures of the limb. Homology refers to features that are shared among different species due to descent from a common ancestor. Image: Understanding Evolution, Figure 7.
Shared features due to a common ancestor are known as homologies, and some can only be viewed in early embryos, whilst others extend into adulthood (see image above). Some homologous structures have no function, like the tail-bone in humans, or the hind leg remnants in whales. These apparently functionless appendages are called vestigial structures, and they can also indicate shared ancestry.
Finally, we have analogous structures: they share function, but not ancestry. An example would be the wings of butterflies and birds, both developed independently as adaptations for similar environmental pressures.
4. Genetics.
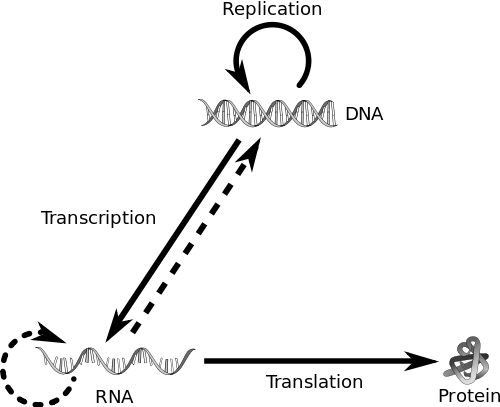 The central dogma of biology. The solid line represents the general case of replication, the dotted line is applicable to certain retroviruses (like HIV). Mutations occur in DNA, but it?s the protein that affects function. Image: Philippe Hupe.
The central dogma of biology. The solid line represents the general case of replication, the dotted line is applicable to certain retroviruses (like HIV). Mutations occur in DNA, but it?s the protein that affects function. Image: Philippe Hupe.
When the discovery of DNA?s double-helix structure was published in 1953, there wasn?t long to wait before the nature of heredity was revealed. DNA was found to be the carrier ? but most importantly, through its own replication, it provided a source of variation.
The building blocks of DNA (adenine, guanine, cytosine, and thymine) are shared throughout the living world, from bacteria to humans. Impressively, triplets of these DNA building blocks form a code that is read to build proteins, and this protein code is also shared throughout life. The result: bacterial DNA can be read in a human cell, and the ultimate indicator for a common ancestor for all life was revealed.
Shared mechanisms aside, sequencing of genomes (all a species? DNA), and the use of mitochondrial ?molecular clocks?, confirmed relatedness theories. We humans share around 99% of our DNA with our closest relative, the chimpanzee, and probably split from our common ancestor a few million years ago. Phylogenies, like the one shown below, can be built to map the genetic differences between species, and so indicate relatedness.
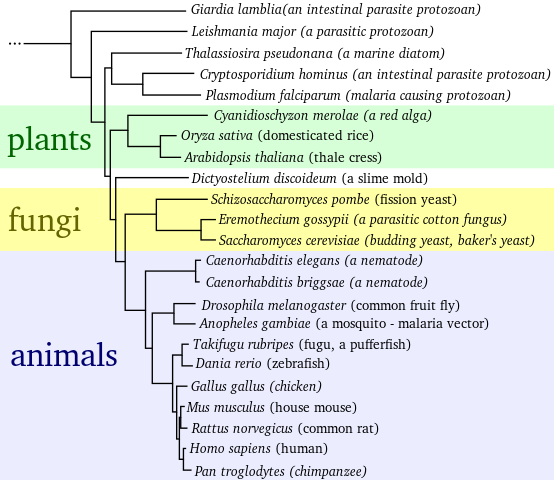 Phylogeny plotting similarity from fully sequenced genomes. Image: Madprime.
Phylogeny plotting similarity from fully sequenced genomes. Image: Madprime.
Variation is generated through mutations in the DNA. This can occur through a variety of mechanisms, such as UV radiation, but most commonly from errors during replication. These errors change how the DNA is read, which changes the protein product and ultimately the function of the cell. Accumulated changes can be positive (new function), neutral or deadly (cancer), and natural selection ultimately removes changes that are negative relative to the local environment.
Which brings us to our final reason.
5. Observable evolution on small timescales.
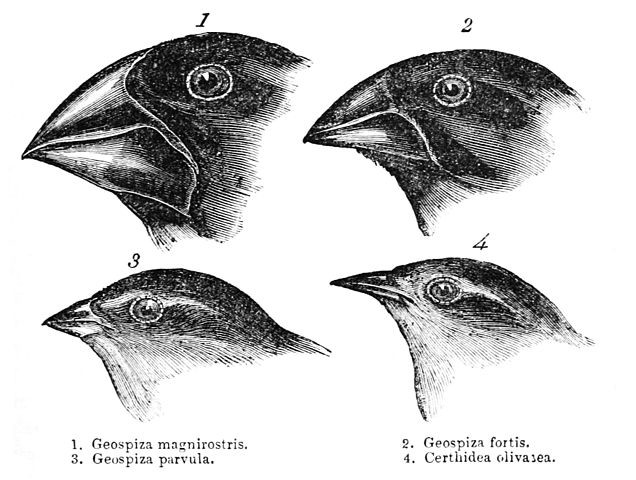 Darwin?s finches from the Galapagos Islands. Each finch shows beak adaptations for a different food source. Image: John Gould.
Darwin?s finches from the Galapagos Islands. Each finch shows beak adaptations for a different food source. Image: John Gould.
Perhaps the most famous of Darwin?s examples of adaptation are the Galapagos finches. Having colonised the volcanic islands from the American mainland, the finches appeared to have different beaks, even though they were similar in all other respects (see image above). It was only much later, after consulting with the ornithologist John Gould, that it was realised that the beaks were adaptations for different food sources. In true natural selection fashion, individuals with better beaks (be it for insects, tough seeds or sucking blood) grew to dominate their local populations. Those who didn?t would slowly die off.
This phenomenon was observed in the Galapagos drought of 2004/5, where average beak size on an island for one species shrank (following mass die-offs of the larger beak variants), likely due to competition for large-beak-associated seeds. The variants with smaller beaks were able to find an alternative food source. Importantly, the genes associated with this trend were identified and their associated increased, or decreased, frequency within the population confirmed.
 The white and black peppered moth. Image: Martinowsky.
The white and black peppered moth. Image: Martinowsky.
Another famous example is the peppered moth, as their population drastically changed following the industrial revolution in England (pictured above).
The expansion of industry in the late 19th century had a major effect on the air quality of British towns, but it also appeared to affect the moth population. The black variation of the peppered moth became increasingly common, while the previously common white variant became rare.
In the 1950s, Bernard Kettlewell showed, through an elegant set of experiments, that the moth population was changing because of predation. The trees surrounding British towns, previously light-barked, had become sooty and dirty in colour, providing better camouflage for the black variant. The white variant, no longer camouflaged, became visible to birds and was hunted in greater numbers. His observations were confirmed in a paper published in 2012; as modern, cleaner forests have seen a reversal back to white variant domination.
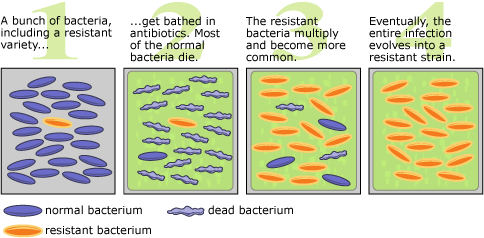 The development of antibiotic resistance through natural selection. Image: University of California Museum of Paleontology?s Understanding Evolution.
The development of antibiotic resistance through natural selection. Image: University of California Museum of Paleontology?s Understanding Evolution.
I couldn?t talk about observable evolution without mentioning bacteria and viruses. The rapid lifecycle of microorganisms means we can observe thousands of generations in a relatively small amount of time.
M. tuberculosis, the bacterial species which causes TB, has been highly efficient at adapting to the selective pressures applied by multiple antibiotics. However, resistance to drugs has developed in various other species of pathogen too. The general mechanism of adaptation to antibiotics is shown in the image above.
Just a theory? Why ?scientific theory? is different.
A ?scientific theory?, in the ideal sense, is the best representation of the available evidence at any one time. The theory itself is not ?fact?, but rather a series of explanations built off observations which are, individually, facts. As such, the theories evolve over time as our understanding of the observations changes, or new observations are made. A scientific theory is always a work in progress.
Of course, there are young, abstract mathematical theories of science, and then there are those which abound with tangible evidence. Evolution, as you?ve seen above, is one of the latter theories.
Currently, there are no observations that have been made which contradict evolutionary theory. It is highly unlikely that there ever will be. But, if some undeniable evidence for a previously unknown mechanism was to suddenly materialize, the scientific community would be forced to revise the affected theories and move on.
Even evolution isn?t infallible; in fact, it wouldn?t be a scientific theory if it was.


{kind=link}
{kind=link}